Research
The molecular genetics of pairing and synapsis
Using super-resolution microscopy to investigate the topology of the SC
The SC is a tripartite structure with lateral elements (LEs) running along the two homologs. The LEs are connected in a zipper-like fashion by pairs of transverse filament proteins which form the central region (CR) of the SC. The N termini of the transverse filaments interact in the middle of the CR and, along with other SC proteins, creates the so-called central element (CE).
The SC is a tripartite structure with lateral elements (LEs) running along the two homologs. The LEs are connected in a zipper-like fashion by pairs of transverse filament proteins which form the central region (CR) of the SC. The N termini of the transverse filaments interact in the middle of the CR and, along with other SC proteins, creates the so-called central element (CE).
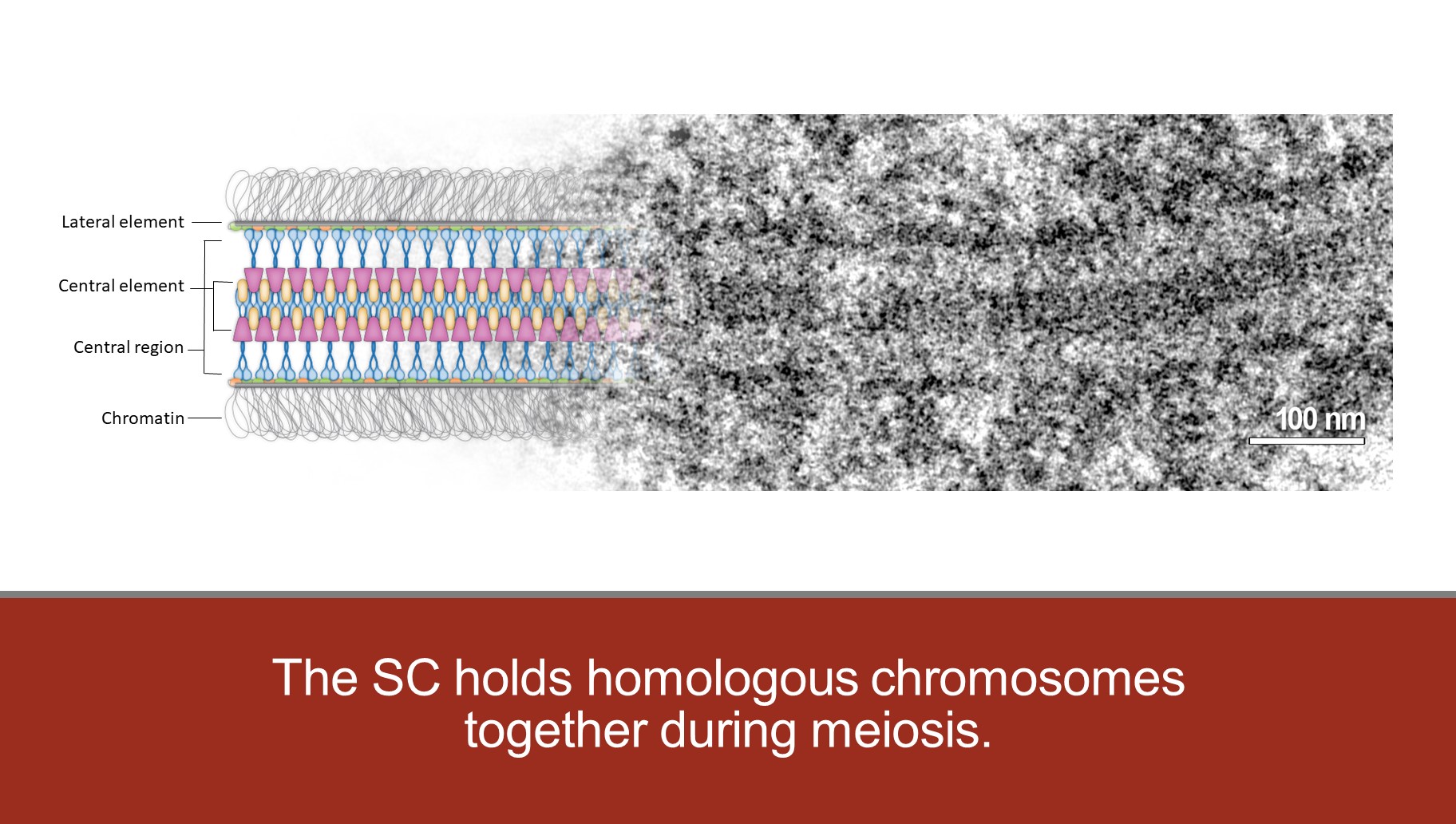

Working with the Stowers Institute Imaging Core, we have combined Structured Illumination Microscopy (SIM) and Expansion Microscopy to provide highly detailed topological maps of SC structure. This study revealed a surprising bi-layered feature of the SC. We are now using this technology to both characterize mutants and further define the position SC components.

The roles of centromere movement in the initiation of meiotic pairing and synapsis
Recent work in our lab and in others’ suggests that the initiation of meiotic pairing and synapsis may be intimately associated with events that involve the centromeres. Moreover, these centromeric movements appear to be connected to the initiation of the assembly of the SC. Our goal is to understand the mechanisms by which synapsis is initiated and the role of centromeres and their flanking heterochromatic regions in this process.
Recent work in our lab and in others’ suggests that the initiation of meiotic pairing and synapsis may be intimately associated with events that involve the centromeres. Moreover, these centromeric movements appear to be connected to the initiation of the assembly of the SC. Our goal is to understand the mechanisms by which synapsis is initiated and the role of centromeres and their flanking heterochromatic regions in this process.
The role of the transverse filament protein C(3)G in pairing and synapsis
Transverse filament proteins are key components of the SC, which connects paired homologous chromosomes during meiosis, ands aids in facilitating recombination. C(3)G, the Drosophila transverse filament protein, spans the width of the SC and is also involved in the early pairing of homologous centromeres. We are performing site-directed mutagenesis to identify critical C(3)G residues, which will allow us to further investigate the timing of SC assembly and maintenance, the nature of the interaction between C(3)G and other SC proteins, and the role of C(3)G in pairing, synapsis, and recombination.
Transverse filament proteins are key components of the SC, which connects paired homologous chromosomes during meiosis, ands aids in facilitating recombination. C(3)G, the Drosophila transverse filament protein, spans the width of the SC and is also involved in the early pairing of homologous centromeres. We are performing site-directed mutagenesis to identify critical C(3)G residues, which will allow us to further investigate the timing of SC assembly and maintenance, the nature of the interaction between C(3)G and other SC proteins, and the role of C(3)G in pairing, synapsis, and recombination.
Understanding the consequences of failed synapsis for crossover positioning
We have recently shown that in-frame deletions within the C(3)G protein create partial loss-of-function mutants that alter the maintenance of synapsis in a polar fashion and have regional and chromosomal effects on crossover frequency. The X chromosome is especially sensitive to such crossover reductions. We are now attempting to more precisely correlate the synapsis and crossover defects and understand the regional specificities we have observed.
We have recently shown that in-frame deletions within the C(3)G protein create partial loss-of-function mutants that alter the maintenance of synapsis in a polar fashion and have regional and chromosomal effects on crossover frequency. The X chromosome is especially sensitive to such crossover reductions. We are now attempting to more precisely correlate the synapsis and crossover defects and understand the regional specificities we have observed.
Identifying components of the central region of the SC
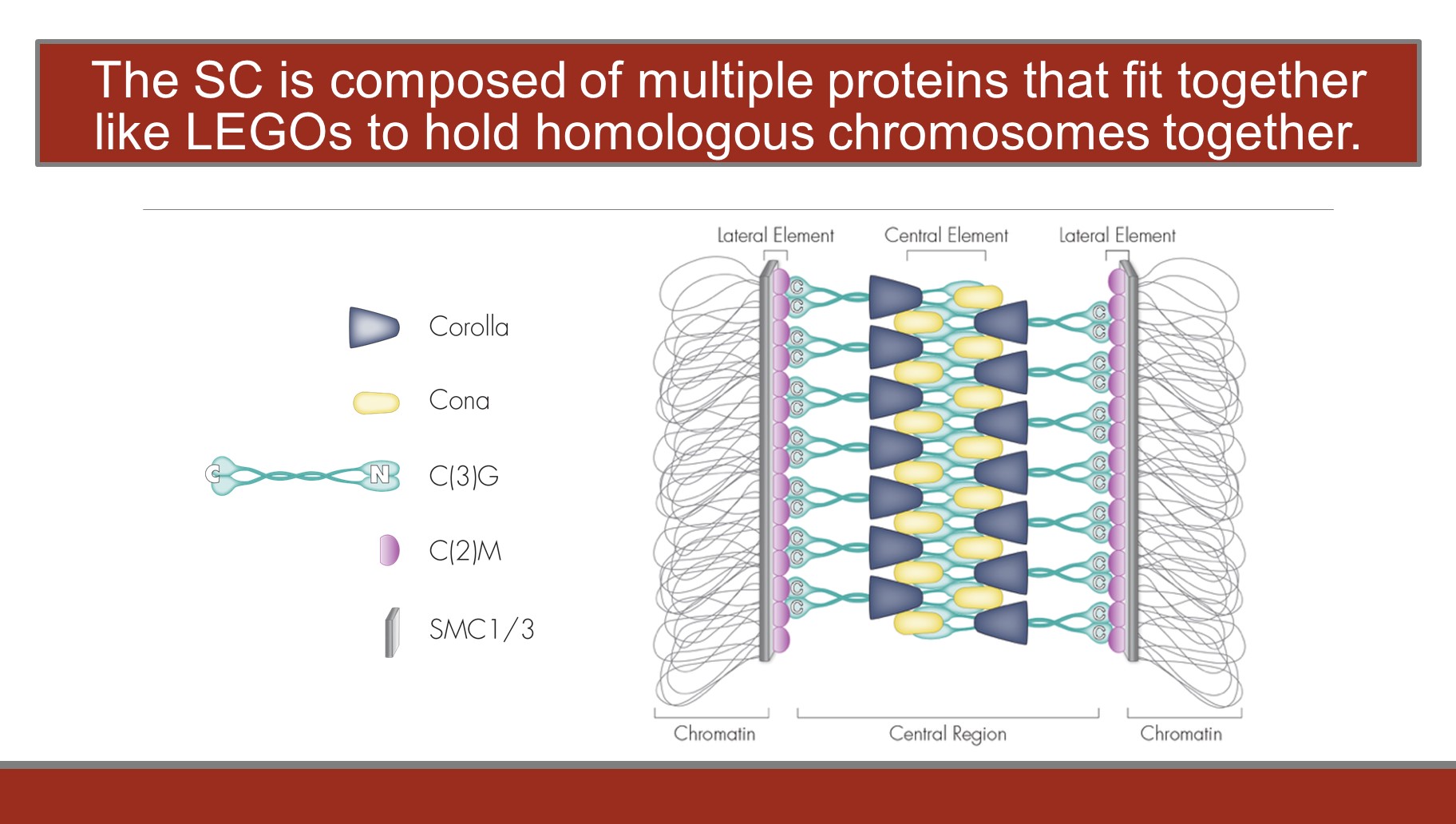
In the last decade we have identified and characterized many new components of the SC. The corolla gene was identified in a screen to isolate novel SC components. corrola mutants are unable to build full-length SC. Similar to other SC mutants, corolla mutants exhibit defective centromere clustering and a severe defect in recombination. Using structured illumination microscopy, we determined that Corolla is a component of the central region of the SC, and a yeast two-hybrid assay indicates that Corolla strongly interacts with Cona. This demonstrates the first direct interaction between two inner-SC proteins in Drosophila.
We also identified Cona a protein that localizes to the central region of the SC. Cona appears to be required to bind transverse filaments together. Cona appears to act as a ‘pillar’ of the central element. We are using multiple approaches to determine just how Cona interacts with the transverse filaments and with other central element proteins (e.g. Corolla). These approaches include a structure/function analysis of the Cona protein as well as genetic and biochemical screens for Cona interactors. Additional screens, both traditional genetic screens and RNAi screens, are underway to identify additional SC components.
In the last decade we have identified and characterized many new components of the SC. The corolla gene was identified in a screen to isolate novel SC components. corrola mutants are unable to build full-length SC. Similar to other SC mutants, corolla mutants exhibit defective centromere clustering and a severe defect in recombination. Using structured illumination microscopy, we determined that Corolla is a component of the central region of the SC, and a yeast two-hybrid assay indicates that Corolla strongly interacts with Cona. This demonstrates the first direct interaction between two inner-SC proteins in Drosophila.
We also identified Cona a protein that localizes to the central region of the SC. Cona appears to be required to bind transverse filaments together. Cona appears to act as a ‘pillar’ of the central element. We are using multiple approaches to determine just how Cona interacts with the transverse filaments and with other central element proteins (e.g. Corolla). These approaches include a structure/function analysis of the Cona protein as well as genetic and biochemical screens for Cona interactors. Additional screens, both traditional genetic screens and RNAi screens, are underway to identify additional SC components.
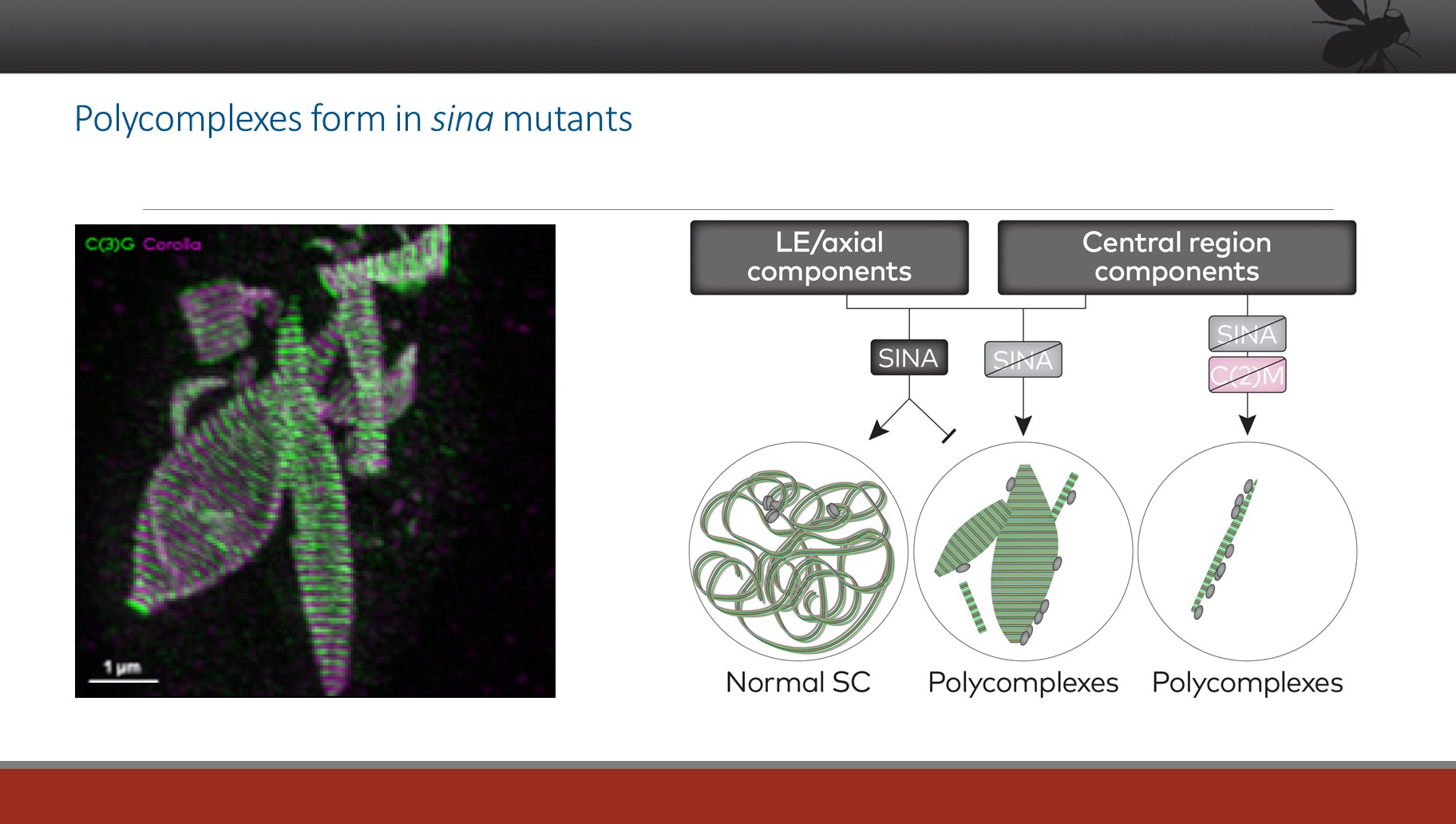
Understanding abnormal patterns of SC assembly
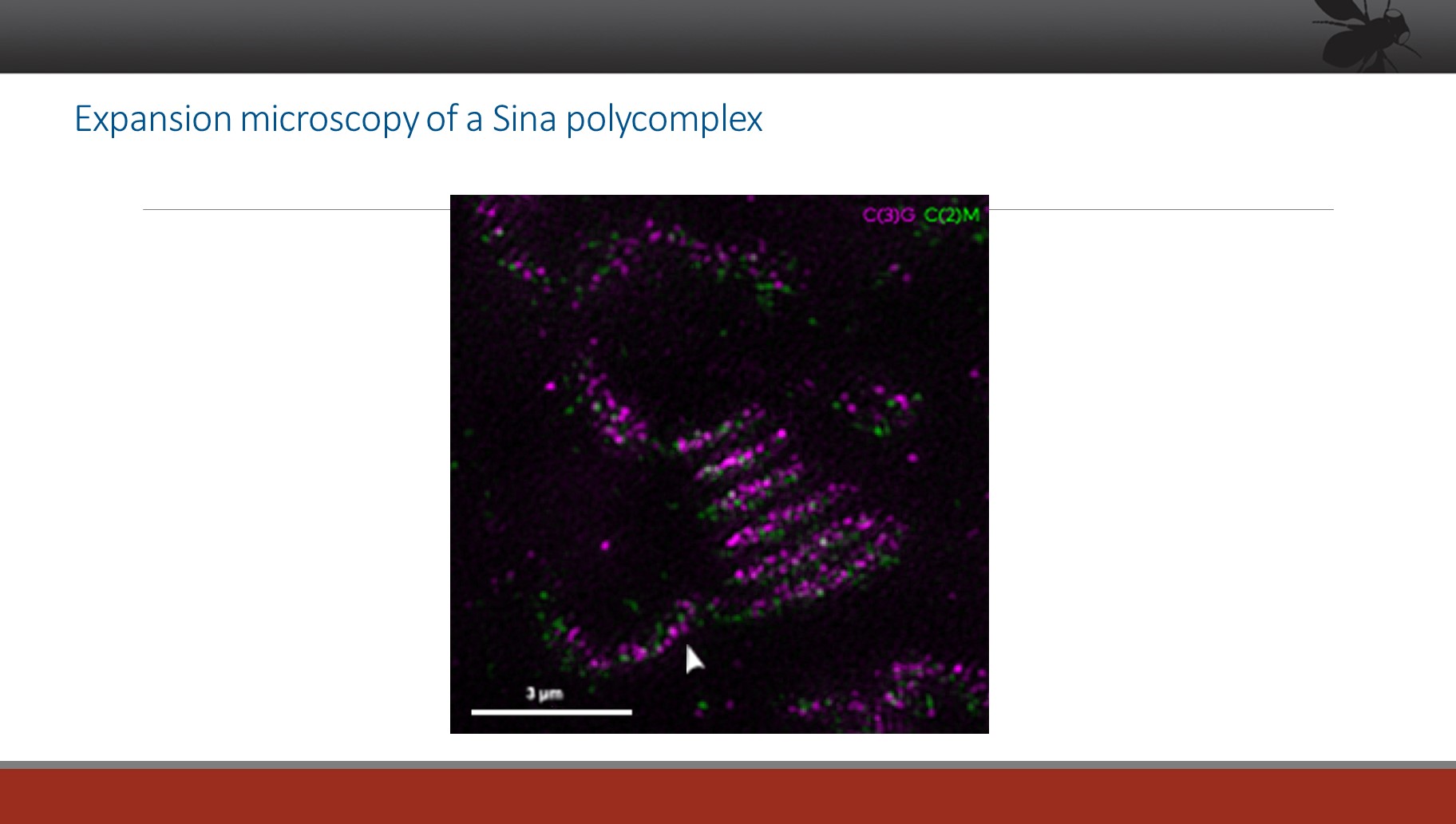
Our efforts to understand the structure of the SC and its assembly process have included a detailed study of a mutant call sina, which allows the formation of grossly aberrant SC structures called polycomplexes (PCs) (see the images below). By understanding how the assembly of such structures is allowed (or induced), we can begin to dissect the pathways that control normal SC assembly.
The molecular biology of the Recombinational Nodule
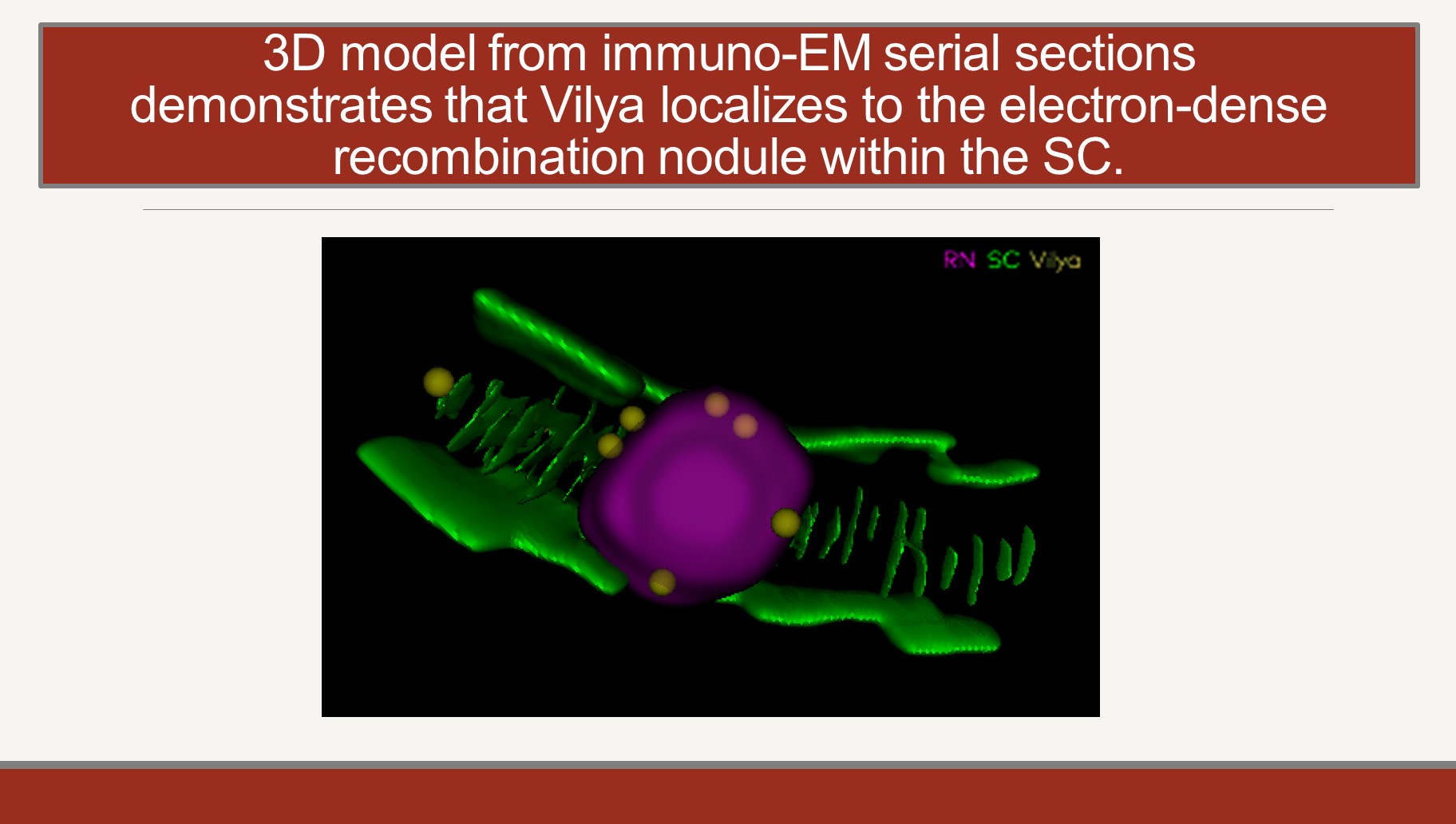
The maturation of crossovers, which requires the breaking and repair of chromosome axes and of two non-sister chromatids, occurs in the context of nuclear structures known as recombination nodules (RN). RNs, which until recently have been studied only by EM, are assembled onto the SC at sites of crossing over. We have identified two components of the RN in Drosophila, the Vilya and Narya proteins. Antibodies against these proteins allow us to study the structure and function of the RN by high resolution light microsopy. We are focused on understanding the function of these proteins in mediating crossover formation and on identifying the proteins with which they physically interact. The short-term goal here is to produce a functional topology of the RN. Our long-term goal is to understand the mechanism by which two chromatids (not just two DNA molecules) can create a crossover.
The maturation of crossovers, which requires the breaking and repair of chromosome axes and of two non-sister chromatids, occurs in the context of nuclear structures known as recombination nodules (RN). RNs, which until recently have been studied only by EM, are assembled onto the SC at sites of crossing over. We have identified two components of the RN in Drosophila, the Vilya and Narya proteins. Antibodies against these proteins allow us to study the structure and function of the RN by high resolution light microsopy. We are focused on understanding the function of these proteins in mediating crossover formation and on identifying the proteins with which they physically interact. The short-term goal here is to produce a functional topology of the RN. Our long-term goal is to understand the mechanism by which two chromatids (not just two DNA molecules) can create a crossover.
The genetic and cytological analysis of meiosis in the silkworm
In the more than 180,000 species that comprise Lepidoptera, many, if not all, female meiotic systems function without crossing over. In the best characterized system, the silkworm Bombyx mori, segregation appears to be accomplished by the large expansion of the SC to form a structure called the Modified Synaptonemal Complex (MSC). As originally described by Rasmussen in the 1970s, the SC undergoes a dramatic structural metamorphosis, increasing its width by more than five-fold to create the MSC. Similar structures have been observed in other lepidopteran oocytes. The silkworm MSC can reach 750 nm in width and is stable until at least anaphase II. This structure bisects the first meiotic spindle, ensuring the segregation of homologs, all of which are achiasmate. This expansion is shown below in images create by Fengli Guo in the SIMR imaging group.
In the more than 180,000 species that comprise Lepidoptera, many, if not all, female meiotic systems function without crossing over. In the best characterized system, the silkworm Bombyx mori, segregation appears to be accomplished by the large expansion of the SC to form a structure called the Modified Synaptonemal Complex (MSC). As originally described by Rasmussen in the 1970s, the SC undergoes a dramatic structural metamorphosis, increasing its width by more than five-fold to create the MSC. Similar structures have been observed in other lepidopteran oocytes. The silkworm MSC can reach 750 nm in width and is stable until at least anaphase II. This structure bisects the first meiotic spindle, ensuring the segregation of homologs, all of which are achiasmate. This expansion is shown below in images create by Fengli Guo in the SIMR imaging group.
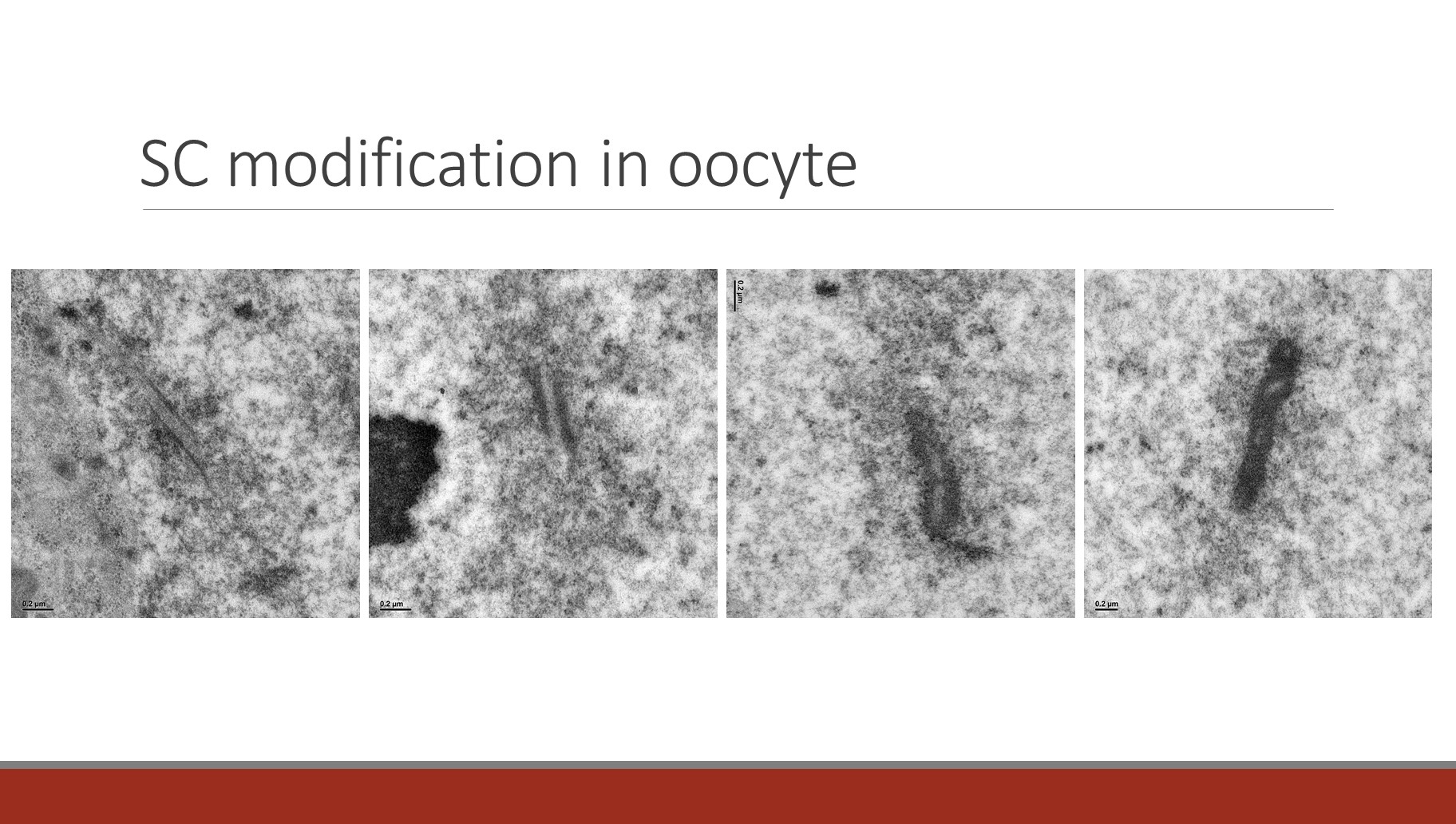
The persistence of the MSC until after homolog separation at Anaphase I appears to be the basis for homolog separation.
We have now identified and characterized the Bombyx orthologs of two key lateral element SC proteins: SYCP3, and HOP1. The search for additional SC orthologs is underway. We are examining the localization of these proteins throughout both oogenesis and spermatogenesis by super resolution microscopy and EM analysis. We will then compare the findings in the achiasmate female system to the apparently canonical meiotic system observed in males. Once key components of MSC formation are identified, models of its structure and formation can be tested by the generation of site-specific mutants by CRISPR. It is at least possible that the large structure and stability of the MSC might make it a reasonable substrate for proteomics analysis.
One last word about teaching and scholarship
Our commitment to meiosis is matched only by our longstanding commitment to teaching. We teach in lecture-based classes and in numerous undergraduate internships in the lab. We view undergraduate, graduate, and postdoctoral training as an essential responsibility to the communities we cherish. We have the same feelings about scholarship; we write books, reviews, and mini reviews as a way to participate in the constant discussion with other scholars with the focus on discovery and education. We are aware how fortunate we are to be able to work with extraordinary colleagues.
We have now identified and characterized the Bombyx orthologs of two key lateral element SC proteins: SYCP3, and HOP1. The search for additional SC orthologs is underway. We are examining the localization of these proteins throughout both oogenesis and spermatogenesis by super resolution microscopy and EM analysis. We will then compare the findings in the achiasmate female system to the apparently canonical meiotic system observed in males. Once key components of MSC formation are identified, models of its structure and formation can be tested by the generation of site-specific mutants by CRISPR. It is at least possible that the large structure and stability of the MSC might make it a reasonable substrate for proteomics analysis.
One last word about teaching and scholarship
Our commitment to meiosis is matched only by our longstanding commitment to teaching. We teach in lecture-based classes and in numerous undergraduate internships in the lab. We view undergraduate, graduate, and postdoctoral training as an essential responsibility to the communities we cherish. We have the same feelings about scholarship; we write books, reviews, and mini reviews as a way to participate in the constant discussion with other scholars with the focus on discovery and education. We are aware how fortunate we are to be able to work with extraordinary colleagues.
References
Combined expansion microscopy with structured illumination microscopy for analyzing protein complexes.
Wang Y, Yu Z, Cahoon CK, Parmely T, Thomas N, Unruh JR, Slaughter BD, Hawley RS. Nat Protoc. 2018 Aug;13(8):1869-1895. doi:10.1038/s41596-018-0023-8. Epub 2018 Aug 2. PMID:30072723
Wang Y, Yu Z, Cahoon CK, Parmely T, Thomas N, Unruh JR, Slaughter BD, Hawley RS. Nat Protoc. 2018 Aug;13(8):1869-1895. doi:10.1038/s41596-018-0023-8. Epub 2018 Aug 2. PMID:30072723
Superresolution expansion microscopy reveals the three-dimensional organization of the Drosophila synaptonemal complex.
Cahoon CK, Yu Z, Wang Y, Guo F, Unruh JR, Slaughter BD, Hawley RS. Proc Natl Acad Sci U S A. 2017;114:E6857-E6866.
Original Data
The E3 ubiquitin ligase SINA regulates the assembly and disassembly of the synaptonemal complex in Drosophila females
Hughes SE, Hemenway E, Guo F, Yi K, Yu Z, Hawley RS. PLoS Genet. 2019;15:e1008161. doi: 10.1371/journal.pgen.1008161.
Original Data
Hughes SE, Hemenway E, Guo F, Yi K, Yu Z, Hawley RS. PLoS Genet. 2019;15:e1008161. doi: 10.1371/journal.pgen.1008161.
Original Data
Vilya, a component of the recombination nodule, is required for meiotic double-strand break formation in Drosophila.
Lake CM, Nielsen RJ, Guo F, Unruh JR, Slaughter BD, Hawley RS. eLife. 2015;4:e08287. doi: 08210.07554/eLife.08287.
Original Data
Lake CM, Nielsen RJ, Guo F, Unruh JR, Slaughter BD, Hawley RS. eLife. 2015;4:e08287. doi: 08210.07554/eLife.08287.
Original Data